

“Daddy, where do babies come from?”
🪢 Sex Without an Ancestor: The Void No Theory Fills

“Sex is the queen of problems... Perhaps no other natural phenomenon has aroused so much interest; certainly none has sowed as much confusion.”
— Graham Bell, biologist.
“The origin and maintenance of sexual reproduction is the most profound mystery in evolutionary biology.”
— George C. Williams, evolutionary biologist.

How did the complex “machinery”
of sexual reproduction arise
“gradually” before it
could be useful?
Evolution is often portrayed as a master of efficiency; a blind sculptor chiselling complexity from simplicity. But then comes sexual reproduction: a system so convoluted, in many ways so costly and counter-intuitive that it defies the very foundations of the principles of natural selection and the “survival of the fittest.”
Sexual reproduction requires two individuals, elaborate signalling, synchronised timing, and the fusion of gametes (ova and sperm) — all for the privilege of producing offspring with only half of the genes. The male provides his genetic pattern without the prospect of bearing offspring. It's like splitting your lottery winnings with a stranger who may or may not show up!
Asexual reproduction, by contrast, is simple, direct, and genetically selfish.
So why would nature abandon it in favour of a system that demands cooperation, vulnerability, and extensive biochemical choreography? Even more profound, though, is the means by which the complex and various attributes of males and females happen to arrive in the first place, in order for synchronised “selection” to take place! Did multiple arrangements arrive fortuitously throughout the enormous “evolutionary timescales” to enable numerous attempts at sexual reproduction?
Even if this is offered as a possibility, it has all the hallmarks of planning and foresight.
The stock evolutionary answer to the question of the arrival of reproduction by sex is “genetic diversity”: sex shuffles the deck, so populations adapt more flexibly. But not only does this violate the stated “goalless mechanism” of evolution, it also presumes the deck was already built; that the shuffling mechanism was already in place, and the benefits were selectable. It's like claiming that the printing press invented itself because people needed books.
The real puzzle is origin: how did the machinery of meiosis, gamete differentiation, and sexual signalling arise before they could be useful? Evolutionists describe this in terms such as “deep evolutionary strategy” without offering any valid evidence or even a reasonable explanation. And strategy requires intelligence, knowledge, foresight, planning... Evolution clearly does not possess these!
Sexual reproduction presents an extraordinary web of interdependencies. Every component relies on each of the others already being in place: no sperm without eggs, no eggs without meiosis, no meiosis without recombination machinery, no recombination without proteins, no proteins without enzymes, in fact no life at all without proteins (see the article 'The Symphony Before the Composition'). The system is not a linear thread but a tightly entangled network of mutually dependent processes.
This raises profound questions for evolutionary theory: If sexual reproduction requires two distinct and complementary reproductive systems, did these systems evolve along perfectly synchronised parallel tracks over vast timescales? How were the anatomical, physiological, and neurological changes in each sex kept precisely aligned at every stage, for countless generations, so that successful reproduction remained possible?
[Technical note:] The improbability of two independently evolving genomic lines arriving at mutual compatibility by chance alone is beyond any practical possibility: the probability that two 3‑billion‑base sequences would match codon-for-codon is in the order of … a one divided by a ten followed by almost 2 billion zeros. In other words, randomness cannot even begin to gesture toward such an outcome.
And note in particular that these “parallel tracks” refer to multiple branches of Darwin's “Tree of Life” — therefore the same models of precisely aligned features discussed above were required, not merely to have evolved in piecemeal synchronicity for one life-form, but somehow to be perpetually available in their completion for multiple occurrences of natural selection to act upon!
Such a scenario would require continuous parallel micro‑level compatibility; not just in broad structures such as gonads or gametes, but in the fine‑grained complexity of the molecular choreographies of meiosis, signalling pathways, receptor–ligand interactions (signals that trigger precise biological responses), gene regulation.
This consistent compatibility would need to be repeated for every sexually reproducing species, which means more than 90% of all life on earth.
The challenge for evolutionists, then, is explaining how gradual, independent mutations could accumulate in two distinct sexes in multiple domains of living creatures, while consistently maintaining the precise micro-synchrony required for the system to continue functioning successfully throughout the alleged eons of evolutionary time!
And all of this from a process that does not see, feel, aim, or plan!
Some theorists invoke symbiosis (a mutually beneficial relationship between two different species), or viral co-option (where design features are somehow “borrowed”) or unspecified ancient cell fusions to cover the quantum leap from asexual to sexual reproduction. But these are speculative scaffolds, not empirical bridges.
🔥 1. Evolution’s “co‑option” assumes foresight
This evolutionary device of “co‑option” is not an explanation but rather a narrative patch. It assumes that complex, interdependent systems can be borrowed, repurposed, and re‑wired before they function; and all this without foresight, and without breaking the organism in the process. When applied to the origin of sexual reproduction, the problems multiply: co‑option requires a chain of coordinated, mutually dependent innovations that:
(1) cannot be staged,
(2) will not function half‑built, and
(3) are useless in isolation.
X and Y chromosomes are the sex chromosomes that determine an individual's biological sex, with XX typically indicating female and XY indicating male.
The evolutionist’s co‑option claims that a structure originally used for X was later repurposed for Y. But for sexual reproduction, Y is not a “new use” — it is a system-level architecture involving:
- meiosis
- gamete differentiation
- gamete recognition
- recombination machinery
- error‑checking
- cell‑fusion protocols
- mating-type compatibility
- regulatory timing
- DNA repair pathways
- germline sequestration (germline is the population of cells that ultimately become male and female sperm and egg cells)
None of these components can “wait around” for the others. They only make sense as a completed system.
Co‑option therefore smuggles in a hidden assumption:
that evolution can anticipate future functionality and preserve non-functional intermediates until the whole system is ready.
And this is foresight by another name; as Juliet put it in Shakespeare's Romeo and Juliet, “That which we call a rose, By any other name would smell as sweet.”
🔥 2. Co‑option requires intermediate stages that cannot exist
For co‑option to work, there must be:
- a prior structure,
- with a prior function,
- that can be modified incrementally,
- without losing viability,
- until it becomes an effective function in a new system.
But for sexual reproduction, the intermediates are catastrophic:
- Half‑meiosis is lethal.
- Half‑recombination breaks genomes.
- Half‑gametes cannot fuse.
- Half‑recognition systems cause sterility.
- Half‑mating types collapse population viability.
Co‑option demands a staircase where empirical biology only offers cliffs.
🔥 3. Co‑option ignores the synchronisation problem
Sexual reproduction requires two partners with matching, compatible, simultaneously emerging systems.
Co‑option narratives must therefore explain:
- how both partners co‑opted the same structures,
- in the same direction,
- at the same time,
- with matching molecular interfaces,
- without any guiding intelligence,
- and without breaking the prior reproductive mode.
This is not a tweak, or an “incremental change.” It is a synchronised overhaul of life’s most fundamental process.
Evolutionary literature rarely addresses this because it is structurally impossible to stage.
🔥 4. Co‑option collapses under the “dependency explosion”
Every co‑option event creates new dependencies:
- If meiosis is co‑opted from mitosis, you must explain recombination.
- If recombination is co‑opted from DNA repair, you must explain gamete formation.
- If gametes are co‑opted from somatic cells, you must explain germline protection.
- If germline protection is co‑opted from stress responses, you must explain mating types.
Each “solution” generates three new problems.
This is why co‑option explanations for sex are always vague, always high‑level, and never mechanistic.
🔥 5. Co‑option assumes that broken systems are tolerated
Co‑option requires long periods where a structure is:
- not doing its original job well,
- not yet doing its new job,
- and is still somehow being preserved by natural selection.
But natural selection eliminates non-functional or harmful intermediates.
Sexual reproduction is especially unforgiving:
Any partial system results in sterility,
and sterility is evolutionary death.
Co‑option therefore contradicts the very mechanism that it relies on!
🔥 6. Co‑option is invoked precisely where evidence is weakest
In evolutionary literature, co‑option appears when:
- no plausible stepwise pathway exists
- no intermediates are known
- no selective advantage can be articulated
- no fossil or molecular evidence supports the transition
It functions as a narrative placeholder, not a demonstrated empirical mechanism.
Co‑option is the evolutionary equivalent of “and then a miracle occurred.”
🔥 7. For the origin of sexual reproduction, co‑option becomes another circular argument
The standard narrative is:
- DNA repair pathways were co‑opted for recombination.
- Recombination was co‑opted for meiosis.
- Meiosis was co‑opted for gamete production.
- Gametes were co‑opted for sexual reproduction.
But each step requires the next step to make sense.
It is a staircase that is built from its own roof!
🔥 8. Co‑option cannot explain the arrival of purpose in the system
Sexual reproduction introduces:
- long-term lineage strategy
- genetic mixing
- population-level benefits
- delayed individual payoff
- increased metabolic cost
- reduced reproductive speed
These are not “borrowed functions.”
They are systemic goals!
Co‑option cannot generate goal‑directed architecture without smuggling in foresight.
The truth is, evolutionists don't know how sexual reproduction began; and their theories, for all their explanatory bravado, quietly sidestep the question!
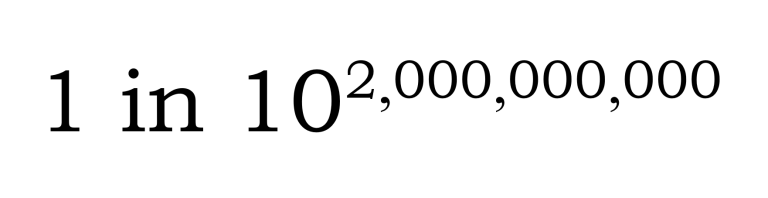
... All of this from a process that
does not see, feel, aim, or plan!

Any partial system results in sterility;
and sterility is evolutionary death.
How likely is it that the 3‑billion sequenced DNA codons in both men and women would match by accident?
